Resumen
Tradicionalmente se ha creído que el sentido del gusto no es importante en las aves ya que estas no son “demasiado exigentes” para el sabor de los alimentos o del agua y poseen una cantidad de receptores gustativos muy inferior a los mamíferos. En campo es común escuchar que los pollos o las gallinas no distinguen los sabores y no tienen sentido del gusto, lo cual es totalmente erróneo. Comercialmente, es frecuente encontrar quejas y reclamos por bajos consumos de alimento en las aves debido al sabor amargo del alimento o del agua. La regulación del consumo está afectada por múltiples factores como el consumo de agua, color, textura, aspectos nutricionales, estado reproductivo, edad, tamaño de partícula y sabor del alimento. Las papilas gustativas juegan un papel clave ya que son las encargadas de detectar cuatro sabores como el amargo, agrio, salado y umami cada una con receptores específicos. El control del apetito en aves es bastante complejo e involucra factores sensoriales en el que participan los principales sistemas de los sentidos como la vista, oído, tacto, olfato y gusto. Esta revisión tiene como objetivo explicar los principales aspectos de la fisiología de la regulación del gusto en las aves y los factores que la afectan, en especial pollo de engorde y ponedoras.

Palabras Claves: Papilas gustativas, Corpúsculos gustativos, Células intragemales, Células gustativas tipo II, Células basales tipo IV, Células perigemales
Introducción
El sentido del gusto (SG) o la percepción del gusto, es la capacidad de evaluar el valor nutricional y potencial tóxico del alimento y convertirlo en un estímulo fisiológico el cual promueve o suprime el consumo de alimento en respuesta a dicho estímulo (Shira et al, 2022). El SG detecta sustancias químicas procedentes de la dieta, las cuales permiten seleccionar dentro de una amplia gama de alimentos, las que son necesarias y beneficiosas para el buen funcionamiento del organismo (Fuentes et al, 2010) y así satisfacer los requerimientos energéticos del ave (Richards, 2003). Además, el sabor permite guiar elecciones nutritivas que motivan la ingesta de alimentos (Xiang Liu et al, 2018).
El SG, está compuesto por las papilas gustativas las cuales están distribuidos en toda la cavidad bucal, y se encargan de detectar moléculas no volátiles a través de receptores del gusto que se expresan en las células sensoriales y que provocan y transmiten impulsos eléctricos hacia el sistema nervioso central para transmitir la sensación del gusto (Shira et al, 2022).
En términos evolutivos, los animales han usado el sentido del gusto para determinar sí el alimento contiene el valor nutricional requerido y sí genera una sensación de placer o hedónica (Shira et al, 2022). Lo anterior, les permite aprender a reconocer y discernir entre alimentos beneficiosos o nocivos (Janssen et al, 2011; Schwartz et al, 2011; Smeets et al, 2012). Por ejemplo, una mala sensación será indicativo de una sustancia potencialmente dañina, mientras que un buen sabor indica comida digestible lo cual es importante para asegurar un consumo óptimo de nutrientes y motivar la ingesta de alimentos adecuada para cubrir los requerimientos (Kare and Herbert, 1960).

en las aves (Te Pas et al, 2020), a pesar de que en la actualidad existan animales con selección genética por caracteres como avidez por consumo de alimento (Torres-Novoa, 2018), y que proporcionan mayor productividad (Valdiviezo, 2012), un bajo consumo de alimento podría incurrir en desbalances energéticos, que no le permitirán expresar su potencial genético ocasionando un bajo desempeño (Bohórquez, 2014).
Existen una variedad de factores que regulan el comportamiento alimenticio, que estimulan o deprimen el apetito (Ferket and Gernat, 2006) como el color (Capretta, 1969; Gentile, 1985), textura (Nir, 1991), aspectos nutricionales (Forbes and Shariatmadari, 1994), temperatura ambiental (Dale et al, 1978; He, 2018), estado del ciclo reproductivo (Savory, 1977; Mench, 2002), edad (Bigot et al, 2003), tamaño de partícula (Nir et al, 1994; Abdollahi et al, 2018) y sabor del alimento (Kare and Herbert, 1960).
Tradicionalmente se ha creído que el sentido del gusto no es importante en las aves ya que estas no son demasiado exigentes para el sabor de los alimentos o del agua. Sin embargo, en campo es frecuente encontrar quejas y reclamos por bajos consumos en las aves debido al sabor del alimento en especial con medicaciones en agua como por ejemplo con tiamulina la cual dismunye el consumo de agua y alimento (Schuhmacher et al, 2006).
Biología del gusto
El gusto favorece la detección de sabores en los alimentos, su función principal es facilitar un primer y rápido análisis de la composición química de sustancias que van a ser ingeridas (Bachmanov and Beauchamp, 2007). Para cumplir dicha función posee unas estructuras especificas llamadas órganos sensoriales, que son sensibles a diferentes estímulos y se encargan de obtener información del entorno (Fortich, 2013).
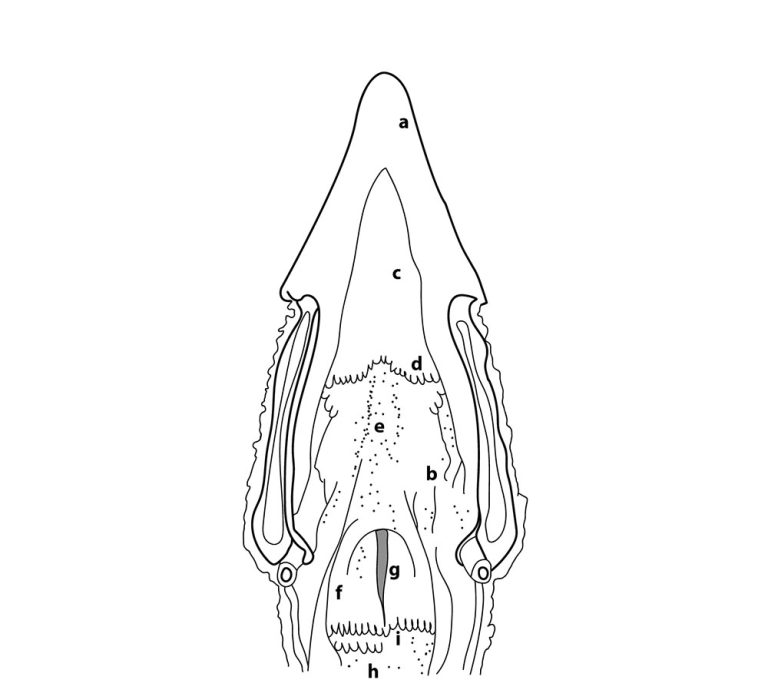
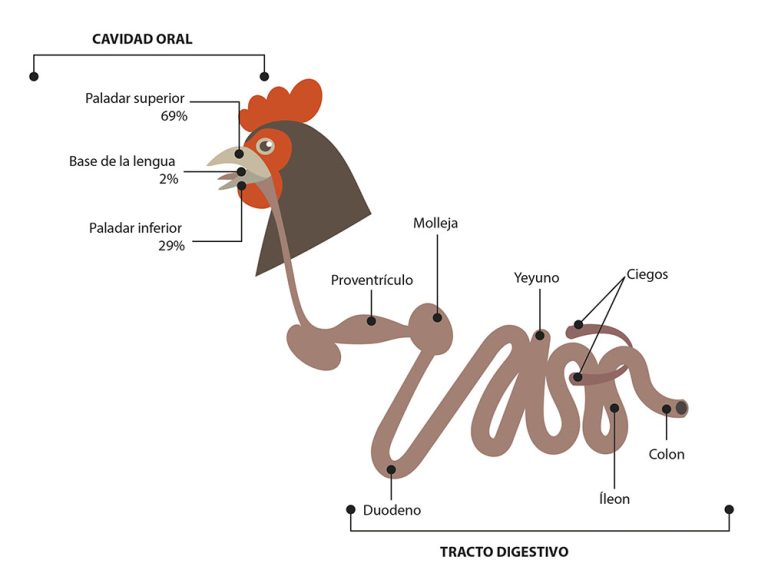
Los órganos sensoriales del gusto son las papilas gustativas (PG), un conjunto de células y receptores sensoriales, encargados de detectar diferentes tipos de sustancias y traducir los estímulos gustativos, por medio de señales neuronales para la percepción del gusto (Roper and Chaudhari, 2017). Están distribuidas y varían en número de manera diferente entre especies animales (Doty, 2015; Davies et al, 1979). En mamíferos, la lengua es el órgano del gusto por excelencia y es allí donde se ubican en mayor medida las PG (Doty, 2015; Travers and Nicklas, 1990). Sin embargo, en las aves esto difiere, ya que se ubican muy cerca de las glándulas salivares, distribuyéndose en las regiones anterior y posterior de la cavidad bucal, específicamente en el paladar (∼69%), la base de la cavidad oral (∼29%), y regiones posteriores de la lengua (∼2%) (Figuras 1 y 2), lo que indica que las aves, a diferencia de los mamíferos, no poseen un sistema gustativo lingual. Dicha distribución estratégica en la cavidad bucal está asociada directamente con el tiempo de contacto del alimento en las distintas zonas, para permitir distinguir mejor los sabores (Berkhoudt, 1977). Además, se pueden encontrar receptores del gusto a lo largo de todo el tracto gastrointestinal. (Figura 3)
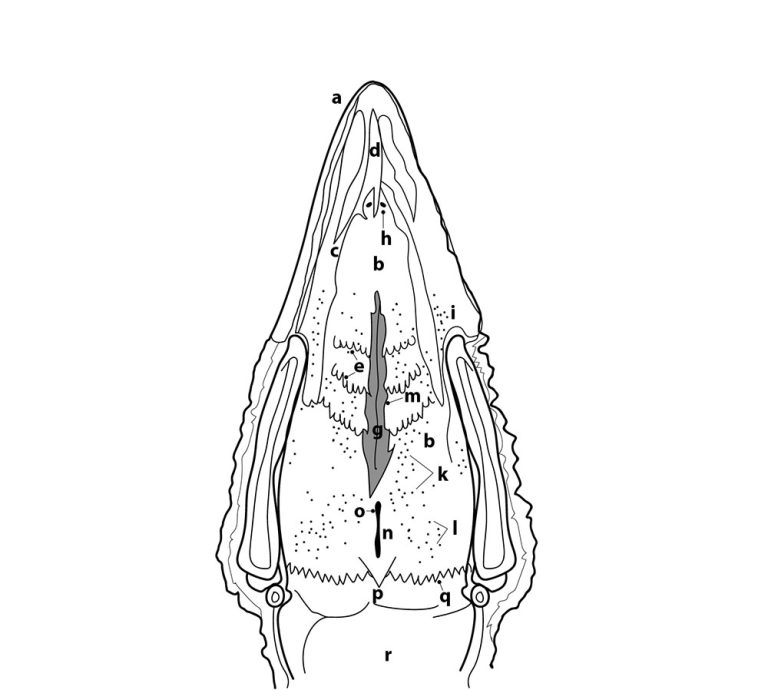
Erróneamente, se creía que las aves no tenían PGs. Sin embargo, hace ya algún tiempo se reportó que tienen entre 360-500 PGs (Ganchrow et al, 1985; Kudo et al, 2010) los cuales varían dependiendo de la edad, especie, raza y sexo e incluso entre línea genéticas (Rajapaksha et al, 2016).

Corpúsculos y células gustativas
Para el correcto funcionamiento de los mecanismos del sentido del gusto es necesario que se active un conjunto de células llamadas células receptoras del gusto las cuales se agrupan en corpúsculos gustativos que forman las PGs con funciones de detectar diferentes modalidades de sabor (dulce, amargo, umami, salado y agrio) y convierten las señales químicas de los alimentos en impulsos eléctricos (Calderón and Arceo, 2020).
En las aves, los corpúsculos gustativos, al igual que en los mamíferos, están compuestas por células especializadas, residentes en epitelio que se alargan y forman una estructura ovoide que rodea el poro gustativo (Calderón and Arceo, 2020). Se han identificado por medio de estudios ultraestructurales, aproximadamente cuatro tipos de células en función de su apariencia, la cual difiere con los mamíferos, aunque con microestructuras similares a las de ellos (DeFazio et al, 2006). Dichas células han sido identificadas por medio de marcadores moleculares utilizados en mamíferos como vimentina y α-gustducina aunque aún no se han reportado dichos marcadores específicos en aves (Kudo et al, 2010; Venkatesan et al, 2016).
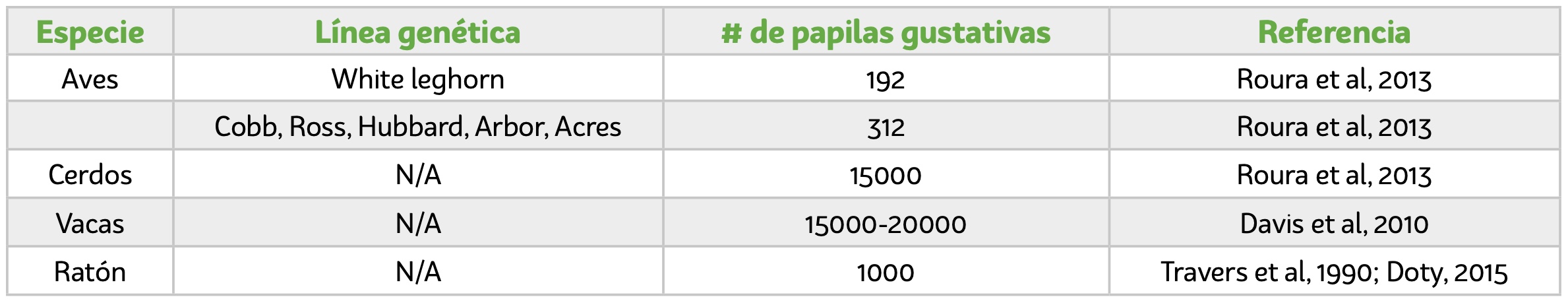
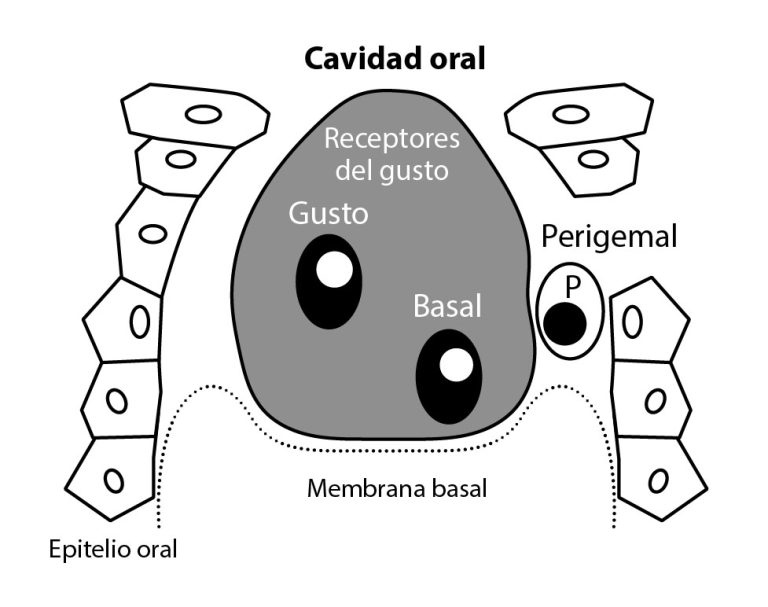
Entre las células descritas para los mamíferos, están las oscuras (tipo I) las cuales son las más abundantes y poseen extensiones citoplasmáticas. Histológicamente, se observan con un citoplasma denso, con cromatina dispersa y algunas vesículas. Su función principal es brindar soporte a la papila (Ganchrow et al, 1991). Otro tipo son las células claras (tipo II), son consideradas células receptoras, tienen citoplasma menos denso y vesículas en mayor número comparada con las oscuras (Sullivan et al, 2010). Las células intermedias (tipo III), poseen unión de características entre los tipo I y tipo II (Figura 4), son de forma aplanada y se encuentran hacia los costados del botón gustativo. Su función principal es la transferencia de señales de neuronas sensoriales (DeFazio et al, 2006; Romanov and Kolesnikov, 2006). Las células basales (tipo IV), están ubicadas en la base de corpúsculo gustativo, tienen un núcleo de forma irregular, con un citoplasma muy denso y se observan más oscuras que las demás. Se denominan células madre o progenitoras, y están involucradas en el recambio celular, el cual dura de 3-4 días (Ganchrow et al, 1994; Ganchrow et al, 1998). Por otra parte, Hong Xiang y colaboradores (2018), encontraron una población de células en proliferación no identificadas anteriormente, que no reaccionaban a los marcadores moleculares convencionales.

Sensibilidad gustativa
La sensación gustativa se produce por el estímulo de receptores mecánicos, térmicos y químicos que están distribuidos por la cavidad oral. Para que se produzca dicho estimulo, el alimento entra en la boca y las sustancias químicas que contienen se disuelven en la saliva y entran en contacto físico con el receptor (mecánicos, térmicos y químicos), estos envían una señal eléctrica al cerebro, la cual finalmente se traduce en el gusto (Miller and Teates 1984; Hyde and Witherley 1993).
Existe una correlación positiva entre el número de receptores gustativos y la sensibilidad al sabor (Berkhoudt, 1985). Por esta razón, se dedujo que en las aves había menor agudeza gustativa, debido al menor número de papilas gustativas comparado con los mamíferos (Roura et al, 2013). Además, las aves poseen menos receptores olfatorios (Doty, 2015).
Esto es importante porque, desde el punto de vista evolutivo, el olfato es un mecanismo quimiosensor utilizado para encontrar fuentes alimento y detectar presas a la distancia (Jones and Roper 1997; Firestein, 2001). Además, detectan moléculas volátiles derivadas de nutrientes esenciales o compuestos potencialmente tóxicos (Pass and Foley 2000; Goff and Klee 2006), por ello, el sentido del gusto y el olfato están estrechamente relacionados, si uno de los dos falla la percepción del sabor cambiará, debido a la estimulación retronasal, esta ocurre durante la ingestión de alimentos (Fuentes et al, 2011). Por medio de la digestión, se liberan moléculas volátiles que son detectadas por el epitelio olfatorio de la nasofaringe, sumado a la detección de los receptores gustativos en la boca hacen parte de la sensación del gusto (Sheperd, 2006; Bartoshuk et al, 2004).
En las aves, los pollos de engorde son más sensibles a estímulos gustativos que los machos de la línea ponedoras (Ganchrow et al, 1985; Kudo et al, 2010), también, los pollos desde el nacimiento responden a bajas concentraciones de estímulos químicos como ácido clorhídrico, ácido acético, entre otros (Gentle, 1972). Además, la sensibilidad gustativa es diferentes para estímulos específicos, los pollos toleran mejor el sabor “agrio” que los mamíferos, pero son muy sensibles al amargo como con el clorhidrato de quinina (Hirose et al, 2015). Responden muy bien a estímulos de glutamato monopotásico que interactúa con el receptor umami (Yoshida et al, 2015). Poseen una respuesta baja al dulce (sacarosa) y al salado (NaCl). Altas concentraciones de NaCl ocasionan rechazo y bajas concentraciones son atractivas, en especial cuando hubo un déficit de sodio. El rechazo inicia en soluciones con 250 mM mientras que la preferencia está entre 85-100 mM (Ganchrow et al, 1990; Gentle, 1972), la calidad del gusto y sensibilidad a los sabores está determinada por la cantidad de receptores gustativos que se explicaran más adelante.
Transducción de señales gustativas y receptores
Se han identificado cinco modalidades o sabores del gusto: dulce, salado, agrio, amargo y umami en mamíferos (Lindemann, 1996; Keast and Breslin et al, 2003). Los pollos pueden detectar solo cuatro de ellos y son el amargo, agrio, salado y umami (Cheled-Shira et al, 2017 ; Yoshida et al, 2018a ; Yoshida et al, 2018b). Cada uno de estas modalidades es reconocido por la PG, pero solo posee una única célula receptora la cual está inervada por fibras nerviosas individuales que transmiten las señales de esa única célula, lo que quiere decir que cada sabor es identificado por un solo tipo de célula.
Los receptores neuroquímicos se clasifican en receptores inotrópicos(iGuR), estos tienen una única proteína receptora específica para un canal iónico especifico, y metabotrópicos, que están asociados a proteína G (Lagerström et al, 2006). Este último es el principal y más relevante, y a este grupo pertenecen los receptores gustativos o receptores TR (taste receptor), los cuales se expresan en las diferentes células que conforman la PG (Fuentes et al, 2010). En las aves se han detectado que los receptores acoplados a proteína G 120, interviene en la respuesta gustativa del ácido oleico y linoleico, y es receptor gustativo de grasas funcionales en los pollos (Sawamura et al, 2015).
Los receptores TR para el dulce (glucosa), amargo y umami (glutamato), pertenecen a una súper familia de receptores acoplados a proteínas de unión a nucleótidos de guanina (GPCR) los cuales se han clasificado en dos grupos T1R y T2R (Muller et al, 2005; Yoshida et al, 2019). En las aves, los T1R se relacionan con umami y dulce, con un receptor específico para umami (T1R1), para dulce (T1R2) y uno para ambos(T1R3) (Yoshida et al, 2015). Las aves solo expresan GPCR T1R (T1R1 y T1R3) que corresponde solo al sabor umami y no poseen codificación para el receptor del dulce(T1R2) (Shi and Zhang 2006; Yoshida et al, 2022). por lo anterior, se puede decir que las aves no pueden saborear el dulce (Roura et al, 2008). El grupo de receptores T2R se definen como receptores del sabor amargo y se han identificado tres funcionales (T2R1, T2R2, T2R7) que son activados por varios agonistas (Behrens et al, 2014; Hirose et al, 2015; Dey et al, 2017), este sabor es sumamente importante en las aves ya que son muy sensibles a dicho sabor, incluso desde el momento del nacimiento (Urata et al, 1992; Cheled-Shira et al, 2017; Dey et al, 2018). La explicación radica en que este tipo de sustancias son las que producen algunos insectos o plantas como mecanismo de defensa (Duffus and Slaugther, 1985; D’mello, 2000), lo que ayuda a las aves a prevenirlas contra la ingestión de dichas sustancias toxicas. Por lo anterior, es importante controlar esos sabores amargos en la formulación de las dietas, ya que los animales podrían limitar su consumo (Muller et al, 2005). Es frecuente encontrar medicaciones solubles en agua las cuales son bastante amargas y generan rechazos tanto en el consumo de agua como de alimento (Lopez and Gutierrez, 2005).
Por otra parte en las bases de datos del genoma del pollo se han identificado genes que codifican receptores para el gusto (Zhao et al, 2015), entre ellos se ha referenciado la presencia del canal de sodio epitelial (ENaC), el cual tiene alta selección hacia el sodio (NaCl) y el principal mecanismo de precepción del sabor salado en las papilas gustativas (Hiyama et al, 2022). En cuanto al agrio, se han expresado el canal PKD2L1 al igual que en los mamíferos (Huang et al, 2006), pero aún no se tienen muchas referencias sobre la detección de este sabor en aves, se relaciona con la acidez de los alimentos en descomposición, por la fermentación bacteriana y, por lo general, provoca una respuesta de rechazo (Bachmanov and Beauchamp, 2007)
Factores limitantes del consumo de alimento

El control de la ingesta involucra mecanismos reguladores basados en la teoría glucostática y la termostática (Gleaves., 1989). La glucostática hace referencia a la glucosa en la sangre y porcentaje de almacenamiento en el hígado, es una vía prioritaria en aves ya que satisface sus requerimientos de energía (Shurlock and Forbes, 1981). También, la teoría termostática la cual consiste en que la temperatura ambiente influye sobre el consumo voluntario asociado con el calor generado por el metabolismo, cuando la temperatura disminuye, promueve el consumo de alimento para generar más calor. opuestamente, cuando la temperatura incrementa, las aves no son capaces de disipar el calor, creando un efecto supresor del apetito (Hurwitz et al, 1980). Para la distensión del tracto gastrointestinal, no se conoce con exactitud el mecanismo por el cual regula el consumo, pero se cree que por medio de receptores dentro del buche se transmiten una señal, generando respuesta de saciedad que detiene el consumo (Rasoamanana et al, 2012).
Por lo anterior se establecen los factores más influyentes en el consumo voluntario del alimento en aves, clasificados en tres grandes grupos: nutricionales, fisiológicos y manejo.
Factores nutricionales (dieta)
Entre los factores más influyentes en la dieta se encuentran: ingredientes de la dieta y porcentaje de nutrientes (Forbes 1998; Richards and Proszkowiec-Weglarz, 2007). En las aves, se ha estudiado los requerimientos esenciales para su mayor desempeño productivo, que dependen de la edad y la etapa productiva, normalmente poseen un alto valor energético y proteico, con bajo valor en fibra, y se basa en un aproximadamente 50% en cereales (maíz), sub productos de molienda(carbohidratos), productos de origen animal (proteínas) y forrajes secos (fibra) (Paulino, 2021).
Uno de los requerimientos principales es la energía. Por ende, es importante ofrecer dietas balanceadas con una alta densidad energética, ya que, si el alimento balanceado no contiene energía suficiente, el animal se ve obligado a aumentar el consumo de carbohidratos hasta suplir sus requerimientos para llegar a la máxima capacidad de almacenamiento en el buche, causando pérdidas de masa corporal y ocasionando pérdidas económicas (Duke et al, 1977; Boorman, 1979).
El contenido de proteína, tiene un efecto más indirecto en la influencia del apetito. Sin embargo, si no se cumple con los requerimientos, ocasionará un retraso en la curva de crecimiento y peso corporal (Garcia et al, 1997). En pollos de engorde, se debe tener un equilibrio con los carbohidratos, ya que una mala formulación puede conllevar a incrementar la conversión alimenticia por baja digestibilidad de los carbohidratos (Tobin and Boorman, 1979).
Las vitaminas y minerales tienen funciones estructurales o como cofactor del metabolismo, su principal efecto sobre el apetito está determinado por deficiencias o excesos (Koutsos et al, 2006). Las deficiencias causan alteraciones en la sensibilidad gustativa, como por ejemplo la deficiencia del zinc afecta la ingesta de agua y mejora las respuestas a los estímulos de sabor amargo y salado en las aves (Beltran et al, 2009). El caso contrario son las deficiencias de vitamina A, las cuales causan disminución de la respuesta a estímulos gustativos, posiblemente por alteraciones de las mucosas (Segura and Boada, 2010), la suplementación con minerales aumenta el consumo de agua (Farfán et al, 2010).

Los factores antinutricionales, son aquellos compuestos que afectan el valor nutricional de algunos alimentos y que por lo general son de origen vegetal. Algunos de estos son los inhibidores de tripsina, goitrógenos, alcaloides, oxalatos y fitatos (Elizalde et al, 2009). Estos dificultan la asimilación de nutrientes y alteran el consumo, por medio de sustancias toxicas o inhibición de enzimas, causando efectos fisiológicos poco deseables como distensión estomacal, afectaciones pancreáticas y aglutinación de glóbulos rojos, entre otros (Shahidi et al., 1997).
Factores fisiológicos (físicos y químicos)
Entre los factores fisiológicos se encuentran el tiempo de digestión del alimento, ritmos de alimentación y factores sensoriales (Ferket and Gernat, 2006).
Las aves requieren alimentos altamente digestibles, ya que la velocidad de tránsito intestinal es alta. El tiempo para la excreción máxima, varía dependiendo del tipo de dieta suministrada y el tamaño de partícula (Gallardo, 1980; Nir et al, 1994). Se ha determinado un tiempo de tránsito intestinal aproximado de 8 horas después de la ingesta de la dieta de fase sólida en pollo de engorde de 1800g (Shires et al, 1987).
El ritmo de alimentación también constituye un factor de trastorno del consumo, ya que instintivamente ellas consumen el alimento entre las horas de luz (aproximadamente durante 12 h). Someter a las aves a más o menos horas de luz u oscuridad, pasarán por proceso de adaptación, y se acostumbrarán a consumir la ración en periodos de tiempo determinados dependiendo del objetivo de la producción (Ferket and Gernat, 2006).
En las aves, el control del apetito está regulado por un conjunto de factores sensoriales en el que participan los principales sistemas sensoriales como la vista, oído, tacto, olfato y gusto (Hess, 1956). La percepción del sabor se da exclusivamente en la cavidad oronasal, donde se encuentran receptores mecánicos, térmicos y químicos, encargados de la percepción del sabor (Matsunami and Amrein, 2003). En conjunto formarán un sistema quimio-sensorial, encargado de recibir información de los diferentes sistemas sensoriales y guiar las elecciones nutricionales asociadas a experiencias previas positivas o negativas (Roura et al, 2008).
En el reconocimiento de los alimentos, participa activamente el sentido de la visión (Çadirciş, 2014). Investigaciones previas han demostrado que, en las aves, al momento del nacimiento, ya tienen las papilas gustativas desarrolladas y funcionales (Sneddon et al, 1998; Ganchrow et al, 1990; Ganchrow et al, 1987; Gentle, 1972). Además de una preferencia innata por alimentos con cierta forma y color, se ha evidenciado preferencias por alimentos de color verde sobre el color rojo (Hess, 1956). En aves adultas, estímulos visión-tacto-gusto, les permite distinguir potenciales fuentes de alimento y reaccionan negativamente a cambios en la dieta (Ferket and Gernat, 2006). Si tienen diferentes tamaños, formas y texturas, ellas se guían por experiencias de aprendizajes previas, más aún si estas elecciones pasadas causaron algún malestar o sensación de disgusto después de consumirlas (Quishpe, 2006).
Factores fisiológicos (físicos y químicos)
Las practicas utilizadas durante el manejo de las explotaciones avícolas, también son un factor determinante sobre el consumo (Ferket and Gernat, 2006). La disponibilidad de agua y comida, diseño y cantidad de comederos y bebederos disponibles, evitará desperdicios de alimento, garantizando la distribución homogénea del alimento evitando competencia entre las aves, y variabilidad en los pesos corporales (Quishpe, 2006). Por otro lado, es importante disminuir factores ambientales (calor, calidad del aire, calidad de la cama), que desencadenen estrés crónico (inmunosupresión) y enfermedad (Widowski, 2010).
Es importante mencionar que el uso de vacunas o presencia de patógenos, que desencadenen una respuesta inmunitaria, causando estrés inmunológico e interfiriendo en la cascada de citoquinas pro inflamatorias (IL-1, IL-6, TNF-a, etc) que modulan directamente el comportamiento alimenticio del ave, presentarán picos febriles y reducirán el consumo de alimento causando un efecto negativo sobre los parámetros productivos de las aves (Koutsos y Klassing, 2001).
Referencias Bibliográficas
- Abdollahi, M. R., F. Zaefarian, and V. Ravindran. Feed intake response of broilers: Impact of feed processing. Animal Feed Science and Technology 237 (2018): 154-165.
- Berkhoudt H. Special sense organs: structure and function of avian taste receptors. Form and function in birds. 1985; 3:463–496.
- Bachmanov, Alexander A., and Gary K. Beauchamp. “Taste receptor genes.” Annual review of nutrition 27 (2007): 389.
- Bartoshuk LM, Duffy BV, Chapo KA, Fast K, Yiee HJ, Hoffman JH, et al. From psychophysics to the clinic: missteps and advances. Food Qual Preference 2004; 15: 617-32.
- Behrens, M., S. I. Korsching, and W. Meyerhof. 2014. “Tuning Properties of Avian and Frog Bitter Taste Receptors Dynamically Fit Gene Repertoire Sizes.” Molecular Biology and Evolution 31 (12): 3216–3227. doi:https://doi.org/10.1093/molbev/msu254.
- Beltrán, Ana L., and Blas N. Cabrera. “Efecto de Mintrex® Zn en combinación con Sulfato de Zn en la producción de pollos de engorde hasta los 42 días de edad.” (2009).
- Bigot, K., Mignon-Grasteau, S., Picard, M., & Tesseraud, S. (2003). Effects of delayed feed intake on body, intestine, and muscle development in neonate broilers. Poultry science, 82(5), 781-788.
- Boggs K, Venkatesan N, Mederacke I, Komatsu Y, Stice S, et al. Contribución de las células subyacentes del tejido conectivo a las papilas gustativas en la lengua y el paladar blando del ratón. Más uno. 2016; 11:e0146475. [PubMed: 26741369
- Bohórquez, A. V. D. (2014). Perspectiva de la producción avícola en Colombia. (Especialización en alta gerencia). Universidad Militar Nueva Granada. Bogotá, Colombia.
- Capretta, PJ, (1969). Animal Beheviour, 17: 229-231
- Çadirci Ş (2014). Prerequisites for Diet Selection in Poultry. Harran Tarım ve Gıda Bilimleri Dergisi, 18(1), 53 – 65
- Calderón, M. L. H., & Arceo, S. D. B. (2020). La bioquímica y fisiología del sabor. Revista de Educación Bioquímica, 38(4), 100-104.
- Cano FG. (2010). Anatomía específica de aves: aspectos funcionales y clínicos. Facultad de veterinaria, Universidad de Murcia, [En línea]. Available: https://www. um. es/anatvet/interactividad/aaves/anatomia-aves-10. pdf.
- Chandrashekar J, Hoon MA, Ryba NJ, Zuker CS. Los receptores y las células del gusto de los mamíferos. Naturaleza. 2006; 444:288–294. [PubMed: 17108952]
- Cheled-Shira, S. L., Reicher, N., Niv, M. Y., & Uni, Z. (2017). Detecting thresholds for bitter, umami, and sweet tastants in broiler chicken using a 2-choice test method. Poultry Science, 96(7), 2206-2218.
- Dale, N. M., & Fuller, H. L. (1978). Effect of ambient temperature and dietary fat on feed preference of broilers. Poultry Science, 57(6), 1635-1640.
- Davies RO, Kare MR, Cagan RH. Distribution of taste buds on fungiform and circumvallate papillae of bovine tongue. Anat Rec. 1979 Nov;195(3):443-6. doi: 10.1002/ar.1091950304. PMID: 507400.
- Dey, B., Kawabata, F., Kawabata, Y., Nishimura, S., & Tabata, S. (2018). Bitter taste sensitivity and the expression of bitter taste receptors at different growth stages of chicks. The Journal of Poultry Science, 0170188.
- Doty, RL. Handbook of olfaction and gustation. 3rd. Wiley Blackwell; Hoboken, New Jersey: 2015. p. 1240
- D’mello, J. Anti-nutritional Factors and Mycotoxins. En: Farm Animal Metabolism and Nutrition. CABI Publishing. Wallingford, Inglaterra, 2000. p.383-403
- Denbow, M.(2000). CHAPTER 12 – Gastrointestinal Anatomy and Physiology, Editor(s): G. Causey Whittow, Sturkie’s Avian Physiology (Fifth Edition), Academic Press, 2000, Pages 299-325, ISBN 9780127476056, https://doi.org/10.1016/B978-012747605-6/50013-4.
- Doty, RL. Handbook of olfaction and gustation. 3rd. Wiley Blackwell; Hoboken, New Jersey: 2015. p. 1240
- DeFazio, RA, Dvoryanchikov, G., Maruyama, Y., Kim, JW, Pereira, E., Roper, SD, & Chaudhari, N. (2006). Separate populations of receptor cells and presynaptic cells in mouse taste buds. Journal of Neuroscience, 26(15), 3971-3980.
- Elizalde, A. D. D., Pismag Portilla, Y. A. M. I. D., & Chaparro, D. C. (2009). Antinutritional factors in eatable seeds. Biotecnología en el Sector Agropecuario y Agroindustrial, 7(1), 45-54.
- Farfán, Charly, Yngrid Oliveros, and Vasco De Basilio. (2010) “Efecto de la adición de minerales en agua o en alimento sobre variables productivas y fisiológicas en pollos de engorde bajo estrés calórico.” Zootecnia tropical 28.3 (2010): 363-373.
- Ferket, PR and Gernat, AG.(2006)- “Factors that affect feed intake of meat birds: a review”. Revista International Journal of Poultry Science 5 (2006): 905-911.
- Firestein, S. 2001. How the olfactory system makes sense of scents. Nature 1413: 211-218.
- Forbes, J. M., & Shariatmadari, F. (1994). Diet selection for protein by poultry. World’s Poultry Science Journal, 50(1), 7-24.
- Fortich, D. L. F. (2013). Instituto superior del profesorado “JV González” departamento de biología programa de biología animal II.
- Fuentes A, Fresno MJ, Santander H, Valenzuela S, Gutierrez MF & Miralles R. (2010). Sensopercepción Gustativa: una Revisión. Int J. Odontostomat., 4(2).161-168, 2010.
- Gallardo, J. 1980. Manejo avícola. Editorial Limusa. México, DF. 645 p.
- Ganchrow D, Ganchrow JR, Royer SM, Dovidpor S, Kinnamon JC. (1998). The proliferation of cells of the taste buds in the chick before hatching. chemical senses. 1998; 23:333–341. [PubMed: 9669046]
- Ganchrow D, Ganchrow JR, Romano R, Kinnamon JC. (1994). Ontogenesis and taste bud cell turnover in the chicken. I. Gemmal cell renewal in the hatchling. J Comp Neurol. 345:105–114. [PubMed: 8089272]
- Ganchrow JR, Ganchrow D, Royer SM, Kinnamon JC. (1993). Aspects of vertebrate gustatory phylogeny: morphology and turnover of chick taste bud cells. Microsc Res Tech. 26:106–119. [PubMed: 8241547]
- Ganchrow JR, Ganchrow D. (1987). Taste bud development in chickens (Gallus gallus domesticus). Anat Rec. 1987; 218:88–93. [PubMed: 3605664]
- Ganchrow JR, Steiner JE, Bartana A. (1990). Behavioral reactions to gustatory stimuli in young chicks (Gallus gallus domesticus). Dev Psychobiol. 1990; 23:103–117. [PubMed: 2365134]
- García R, Villanueva V, Cepeda A. y Padrón E. 1997. NM 25. Comportamiento de pollos bajo restricción alimenticia. Archivos Latinoamericanos Producción Animal 5(Supl. 1): 319-320. Disponible en http://www.alpa.org.ve/PDF/Arch%2005%20Suplemento/NM25.pdf
- Gentle MJ. Taste preference in the chicken (Gallus domesticus L). Br Poult Sci. 1972; 13:141–155. [PubMed: 5017939]
- Gleaves, EW.(1989). Application of feed intake principies to poultry care and management. Poilt.Sci., 68: 958-969
- Goff, S. A., Klee, H. J. (2006). Plant volatile compounds: Sensory cues for health and nutritional value?, Science 311: 815-819
- He, X., Lu, Z., Ma, B., Zhang, L., Li, J., Jiang, Y., … & Gao, F. (2018). Effects of chronic heat exposure on growth performance, intestinal epithelial histology, appetite‐related hormones and genes expression in broilers. Journal of the Science of Food and Agriculture, 98(12), 4471-4478.
- Hidalgo, L. A. S.(2016). Uso de vitaminas en pollos de engorde. [En línea]: https://www.agrovetmarket.com/resources/investigacion_y_desarrollo/articulos_tecnicos/uso-de-vitaminas-en-pollos-de-engorde-213170d71.pdf
- Hirose N, Kawabata Y, Kawabata F, Nishimura S, Tabata S. Bitter taste receptor T2R1 activities were compatible with behavioral sensitivity to bitterness in chickens. Biochem Biophys Res Commun. 2015; 460:464–468. [PubMed: 25796330]
- Hyde, R. J. and Witherly, S. A. 1993. Dynamic contrast: Asensory contribution to palatability. Appetite 21: 116
- Hurwitz, S, Weiselberg M, Eisner U, Bartov G, R iesenfeld, M. Sharvit, A. Niv y S. Bornstein. (1980). The energy requirements and performance of growing chickens and turkeys as affected by environmental temperature. Poult. Sci., 59: 2290-2299.
- Jones, R. B. and Roper, T. J. 1997. Olfaction in the domestic fowl: a critical review. Physiol. Behav. 62: 10091018
- Kare, Morley R., and Herbert L. Pick Jr. “The influence of the sense of taste on feed and fluid consumption.” Poultry Science 39.3 (1960): 697-706. https://doi.org/10.3382/ps.0390697
- Kawabata, Fuminori, and Shoji Tabata. “Bitter Taste Perception in Chickens.” The Journal of Poultry Science (2021): 021
- Koutsos, EA and Klassing KC.(2001). Modulation of nutritional status by the inmune response. In: Proceeding 28th Annual Carolina Poultry Nutrition Conference, pp: 1-6. November 14-15th, Sheraton Imperial Hotel, Raleigh, NC
- Keast, R. S., & Breslin, P. A. (2003). An overview of binary taste–taste interactions. Food quality and preference, 14(2), 111-124.
- Kudo K, Nishimura S, Tabata S. (2008) Distribution of taste buds in layer-type chickens: scanning electron microscopic observations. Anim Sci J. 2008; 79:680–685.
- Kudo K, Wakamatsu K, Nishimura S, Tabata S. (2010). Gustducin is expressed in the taste buds of the chicken. Anim Sci J. 81:666–672. [PubMed: 21108686]
- Lagerström MC, Hellström AR, Gloriam DE, Larsson TP, Schiöth HB, et al. The G protein– coupled receptor subset of the chicken genome. PLoS Comput Biol. 2006; 2:e54. [PubMed: 16741557]
- Lindenmaier P, Kare MR. The taste end-organs of the chicken. Poult Sci J. 1959; 38:545–550.
- Lindemann, B. (1996). Taste reception. Physiological reviews, 76(3), 719-766.
- Liu, HX, Rajapaksha, P., Wang, Z., Kramer, NE y Marshall, BJ (2018). Una actualización sobre el sentido del gusto en los pollos: un sistema mejor desarrollado de lo apreciado anteriormente. Revista de nutrición y ciencias de la alimentación, 8 (2).
- López, Héctor Sumano, and Lilia Gutiérrez Olvera. “Consideraciones Farmacológicas de la Antibióticoterapia em Aves.” Asociación Ecuatoriana de Buiatría (2005).
- Matsunami, Hiroaki, and Hubert Amrein. “Taste and pheromone perception in mammals and flies.” Genome biology 4.7 (2003): 1-9.
- Miller, M. G. and Teates, J. F. 1984. Oral somatosensory factors in dietary self-selection after food deprivation and supplementation. Behav. Neurosci. 98: 424434.0017.
- Mench, J. A. (2002). Broiler breeders: feed restriction and welfare. World’s Poultry Science Journal, 58(1), 23-29.
- Mennella, J. A., Forestell, C. A., Morgan, L. K., & Beauchamp, G. K. (2009). Early milk feeding influences taste acceptance and liking during infancy. The American journal of clinical nutrition, 90(3), 780S-788S.
- Mueller, K. L., Hoon, M. A., Erlenbach, I., Chandrashekar, J., Zuker, C. S., & Ryba, N. J. (2005). The receptors and coding logic for bitter taste. Nature, 434(7030), 225-229.
- Niknafs, S. y Roura, E. (2018). Detección de nutrientes, sabor e ingesta de alimento en especies de aves. Revisiones de investigaciones sobre nutrición, 1–11. doi:10.1017/s0954422418000100
- Nir, I. (1991). Influencia de la textura del pienso en el rendimiento de los pollitos de engorde. Selecciones avícolas, 33(10), 0699-699.
- Nir, I., Twina, Y., Grossman, E., & Nitsan, Z. (1994). Quantitative effects of pelleting on performance, gastrointestinal tract and behaviour of meat‐type chickens. British Poultry Science, 35(4), 589-602.
- Pass, G. J. and Foley, W. J. 2000. Plant secondary metabolites as mammalian feeding deterrents: separating the effects of the taste of salicin from its post-ingestive consequences in the common brushtail possum (Trichosurus vulpecula). J. Comp. Physiol. 170: 185192.
- Paulino JA.(2021). Los requerimientos nutricionales de las aves dependen de varios factores. Artículos técnicos, avicultura. Engormix
- Pérez-Rodríguez, L. (2016). El sentido del gusto de las aves. Estación Biológica de Doñana (CSIC). Recuperado de file. C:/Users/User/Downloads/12-EL% 20SENTIDO, 2.
- Quishpe, G. J. (2006). Factores que afectan el consumo de alimento en pollos de engorde y postura.
- Rajapaksha P, Wang Z, Venkatesan N, Tehrani KF, Payne J, et al. (2016). Labeling and analysis of chicken taste buds using molecular markers in oral epithelial sheets. Sci Rep. 6:37247. [PubMed: 27853250]
- Rasoamanana, R., Darcel, N., Fromentin, G. y Tomé, D. (2012). Detección de nutrientes y señalización por parte del intestino. Actas de la Sociedad de Nutrición, 71 (4), 446-455. doi:10.1017/S0029665112000110
- Richards, M. P. (2003). Genetic regulation of feed intake and energy balance in poultry. Poultry science, 82(6), 907-916.
- Richards MP, Proszkowiec-Weglarz M. (2007). Mechanisms Regulating Feed Intake, Energy Expenditure, and Body Weight in Poultry. Poult Sci 86: 1478-1490.
- Romo, D. P. Los sentidos del gusto y del olfato en las aves. Midia Relaciones, SA de CV, Mexico (Mexico).
- Roper, S. D., & Chaudhari, N. (2017). Taste buds: cells, signals and synapses. Nature Reviews Neuroscience, 18(8), 485-497.
- Roura, E., Humphrey, B., Tedo, G. e Ipharraguerre, I. (2008). Desplegando los códigos de apetito alimentario a corto plazo en animales de granja y de compañía. Una revisión comparativa de la biología de detección de nutrientes oronasal. Revista canadiense de ciencia animal , 88 (4), 535-558.
- Savory, C. J. (1977). Effects of egg production on the pattern of food intake of broiler hens kept in continuous light. British Poultry Science, 18(3), 331-337.
- Sawamura R, Kawabata Y, Kawabata F, Nishimura S, Tabata S. El papel del receptor acoplado a proteína G 120 en la detección de ácidos grasos en tejidos orales de pollo. Biochem Biophys Res Comun. 2015; 458:387–391. [PubMed: 25656577]
- Schuhmacher A, Bafundo KW, Islam KM, Aupperle, H, Glaser, R Schoon, HA, Gropp JM (2006). Tiamulin and Semduramicin: Effects of Simultaneous Administration on Performance and Health of Growing Broiler Chickens. Poultry Science 85:441-445.
- Segura, O.I.; Boada, M.A. (2010). Efecto de suplementación en la dieta con BIG EGG en los parámetros productivos de ponedoras de huevo commercial. Revista Colombiana de Ciencia Animal. Colombia.Vol. 3, Nº 1. 23-32 p.
- Shepherd GM. Smell images and the fl avour system in the human brain. Nature 2006; 444 (7117): 316-21.
- Shira LCS, Zehava U, Colin GS. Taste in Birds. Sturkie’s Avian Physiology Book Academic Press 2022
- Shires, A., Thompson, J. R., Turner, B. V., Kennedy, P. M., and God, YK.(1987). Rate of passage of corn-canola meal and cornOsman, A. M. (1982). Amylase in chicken intestine and pancreas. soybean meal diets through the gastrointestinal tract of broiler Comp. Biochem. Physiol. B 73, 571–574. and White Leghorn chickens. Poult. Sci. 66, 289–298.
- Shurlock, TG and Forbes JM.(1981). Br. Poult Sci., 22: 333-346.
- Simitzis, P. E., Deligeorgis, S. G., Bizelis, J. A., & Fegeros, K. (2008). Feeding preferences in lambs influenced by prenatal flavor exposure. Physiology & behavior, 93(3), 529-536.
- Smeets, P. A., Charbonnier, L., van Meer, F., van der Laan, L. N., & Spetter, M. S. (2012). Food-induced brain responses and eating behaviour. Proceedings of the Nutrition Society, 71(4), 511-520.
- Sneddon, H., Hadden, R., & Hepper, P. G. (1998). Chemosensory learning in the chicken embryo. Physiology & Behavior, 64(2), 133-139.
- Sullivan JM, Borecki AA, Oleskevich S.(2010). Stem and progenitor cell compartments within adult mouse taste buds. Eur J Neurosci.; 31:1549–1560. [PubMed: 20525068]
- Te Pas MFW, Borg R, Buddiger NJH, Wood BJ, Rebel JMJ, van Krimpen MM, Calus MPL, Park JE, Schokker D. Regulating appetite in broilers for improving body and muscle development – A review. J Anim Physiol Anim Nutr (Berl). 2020 Nov;104(6):1819-1834. doi: 10.1111/jpn.13407. Epub 2020 Jun 26. PMID: 32592266; PMCID: PMC7754290.
- Travers, S. P., & Nicklas, K. (1990). Taste bud distribution in the rat pharynx and larynx. The Anatomical Record, 227(3), 373–379. doi:10.1002/ar.1092270313
- Torres, Jefrey G. “Efecto de fuentes lipídicas en el desempeño productivo de pollitas de reemplazo ponedoras Dekalb White®.” (2021).
- Torres, Diana Milena. “Exigencias nutricionales de proteína bruta y energía metabolizable para pollos de engorde.” RIAA 9.1 (2018): 6.
- Urata, K., Manda, M., & Watanabe, S. (1992). Behavioral study on taste responses of hens and female Japanese quails to salty, sour, sweet, bitter and umami solutions. Animal Science and Technology (Japan).
- Valdiviezo, H. M. F. (2012). Determinación y comparación de parámetros productivos en pollos broiler de las líneas Cobb 500 y Ross 308, con y sin restricción alimenticia. (Tesis de grado). Escuela Superior Politécnica del Chimborazo. Riobamba. Ecuador
- Venkatesan N, Rajapaksha P, Payne J, Goodfellow F, Wang Z. ( 2016). Distribution of α-gustducin and vimentin in premature and mature taste buds in chickens. Biochem Biophys Res Commun.; 479:305–311. [PubMed: 27639649]
- Widowski, T. (2010). The Physical Environment and Its Effect on Welfare. In: Duncan, I., Hawkins, P. (eds) The Welfare of Domestic Fowl and Other Captive Birds. Animal Welfare, vol 9. Springer, Dordrecht. https://doi.org/10.1007/978-90-481-3650-6_6
- Xiang Liu H, Rajapaksha P, Wang Z, Kramer NE, Marshall BJ.(2018). An Update on the sense of taste in chickens: A better developed system than previously appreciated. J Nutr Food Sci. 2018 ; 8(2): . doi:10.4172/2155-9600.1000686.
- Yoshida, Y., Wang, Z., Tehrani, K. F., Pendleton, E. G., Tanaka, R., Mortensen, L. J., … & Kawabata, F. (2019). Bitter taste receptor T2R7 and umami taste receptor subunit T1R1 are expressed highly in Vimentin-negative taste bud cells in chickens. Biochemical and biophysical research communications, 511(2), 280-286.
- Yoshida, Y., Nishimura, S., Tabata, S., & Kawabata, F. (2022). Chicken taste receptors and perception: recent advances in our understanding of poultry nutrient-sensing systems. World’s Poultry Science Journal, 78(1), 5-20.
- Zhao, H., Li, J., Zhang, J., 2015. Molecular evidence for the loss of three basic tastes in penguins. Curr. Biol. 25 (4), R141eR142.